| Abstract
The time of the last evening return to nest of a pair of swifts, which successfully raised two chicks under good weather conditions, was registered for the whole period of residence at the nesting site. Times were recorded relative to the time of sunset and both the weather and the overall breeding pattern were taken into consideration . It was noted that the actual time of sunset for swifts is only one factor, albeit an important one, determining the actual moment of evening return. Points of reference are the light intensity, as well as weather conditions and social factors. The importance of social interactions of the parents and chicks has been a major factor in this investigation. It is hoped that this study will provide the impetus for further work to be undertaken in this regard. |
| ULRICH TIGGES:
|
| On the breeding phenology of the Common Swift (Apus apus) - the last diurnal return to the nest with reference to environmental and social factors |
|
1. Introduction Swifts display two different roosting strategies. While non-breeders fly high into the sky at late dusk to spend the night on the wing (Weitnauer 1960), the breeding pairs roost in the nesting sites. They only spend the nights flying when their presence with hatching is no longer essential or if access to the nest is foiled for any reason, (cf. Lack & Lack 1952, Weitnauer 1960, Keller 1977, personal observation; Kumerloeve (1954) reports nocturnal hunting). Rising into the sky as well as descending at dawn has been described and discussed for 200 years (Spallanzani 1797 in Weitnauer & Scherner 1994, Guérin 1923, Weitnauer 1952, 1954, 1960). More general statements about the roosting times have earned numerous references in the literature. They describe the end of the flying patterns in the sky (e.g. Hoffmann 1917, Schuster 1921, Sehlbach 1926, Haartman 1940). Auel 1917, Haartman 1940, Daanje 1944, Haartman 1949, Vleugel 1947, Church 1956 and Arn-Willi 1960 relate the last descent to the position of the sun and cite the number of minutes the swifts stay on the wing beyond sunset. Table
1. The farther north the nesting sites are, and consequently the longer dusk
lasts, the later swifts continue to fly after sunset.
Only Haartman (1949) quoted the brightness of the roosting time in the unit of measurement Lux (cf. note). The values he published are not natural measurements though, but are calculated. This certainly is due to the technical possibilities in his time. As a result of his investigation he stipulated that the swift maintained a certain level of light intensity, no matter what the season, geographical latitude or question of an overcast sky, „corresponding to about 4-15 Lux.“ He presupposes the swift would continue its flight until midnight or beyond if the prevailing light did not diminish into this range. According to his observations the last return to the nest takes place about 10 minutes earlier when skies are overcast, which ought to correspond to the same light intensity as the unclouded sky at this latitude. Technological progress means that anyone today can obtain a light meter which permits us to publish the previously missing data. Furthermore maximum and minimum temperatures as well as additional information during the evening hours can be noted during the period of observation. In addition, biological factors concerning breeding patterns which appeared relevant during this study were noted. By means of these observations it was possible, to investigate behavioral influences on the last diurnal return. 2. Materials and Methods It can be stated with certainty that the breeding pair observed in this study was experienced, since one of the birds had been ringed as early as 1992. The nesting site is 16 meters above ground level. It is in a wooden box under the window sill of a four-flat old building in a central heavily built up area of Berlin. The distance to the opposite row of houses is 20 meters. The top of the row of linden trees does not reach the level of the nest, leaving a free space under the entrance. The pair is breeding solitarily, not under the influence of other breeding swifts in the neighborhood. The next breeding pair is probably located on the rear side of the house facing. Immediately after the second bird entered (apx. 10 - 30 seconds) I measured the light intensity at a location about 2 meters horizontally from and 1 metre below the nest box. I used a standard Digital Lux Meter (according to the manufacturer, the accuracy has been tested by a standard parallel light tungsten lamp of 2854° K temperature; accuracy ± 5% + 2d). To gain comparable measurements I always placed the light sensor at the same point and registered the highest light value there. (For more complete understanding it is useful to know that the Lux is a very small unit. A tiny change in the position of the sensor registers different intensities of lightness. To lower this uncertainty factor I always tried to register the highest light level at the same place.) Daily temperatures were measured with a maximum/minimum thermometer. It was fixed on the door of the balcony about 90 cm parallel to and 40 cm below the nest box. The measured values tend to be about 2° C more moderate than the values published by the authorities, but correspond to those at the nest site. The hours of sunset were provided by the German Meteorological Service and refer to the meteorological office at Tempelhof Airport, which is about 2.7 kilometers due west of the nesting site. The fertilised eggs were laid on May 28th and 30th 1995 which is within the range for this latitude (Litzbarski, in Rutschke 1987). The 1995 weather was cool and rainy until June 24th. From June 27th on a stable period of very warm dry weather started and did not end before the swifts left on their autumn migration. It is well known that swifts feed on air born insects and therefore the conditions of weather were highly favourable for reproduction. On rainy days there was invariably enough time without precipitation to permit the birds to feed. Regular observations of flying activities to and from the nest box took place on the balcony or from the adjacent room. Monitoring was continuous during the expected time for entering the nest in the evenings, correspondingly earlier in times of bad weather. In general I was even able to record the time before the last entering of the second bird in 1995, so that all sensitive phases, such as egg laying or at the end of the breeding period could be registered completely with the exception of five specific days. The last enterings of the first inflying bird were registered 69 times. After watching two attempts to enter another nest box at the rear of the building, at only 1 Lux illumination at 9:45 p.m. on May 17th, I prolonged the time of observing the breeding pair until it was completely dark, despite my experiences from previous years. I observed no further flight activity however. The lightness is of different intensity on just about every point in the area. Because of the buildings the light values just in front of the nest box are definitely lower than several meters higher. The data which are listed here do not attempt to represent the intensity of light even a few feet away from the nesting site. This means that the values of the immediate inflight surroundings are registered and not those which provide signals to the bird in flight to return to their roost. These values must be considerably higher. The bird has to calculate in advance when to go back to the nest to pass through the entrance hole successfully. Mistakes are made but these appear to be of extremely rare occurrence (Weitnauer 1990). 3. Results In the year 1995 the times of the latest entering the nest of a breeding pair of swifts were noted and analyzed together with the intensity of light, weather conditions as well as certain biological data. The result is drawn in Figure 1. The weather conditions had been good so the behavior of the parent birds in this breeding season could be taken as exemplary. The
seasonal sojourn of the swifts in our part of the world can be divided into four
periods which have an effect on the nest entering times:
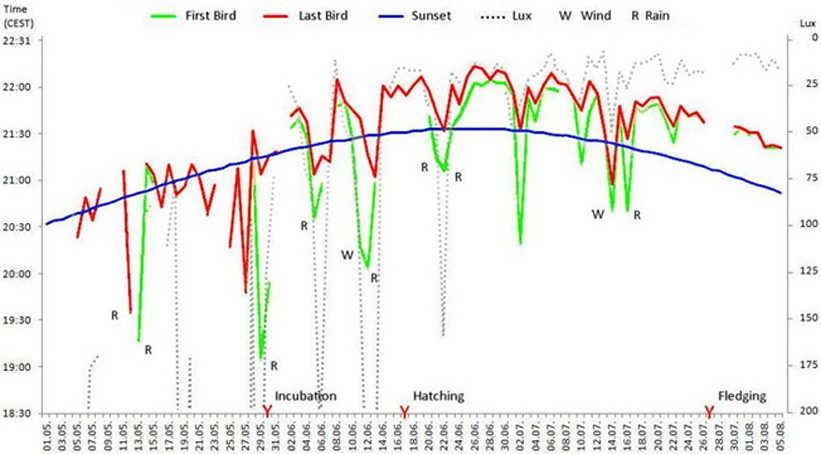
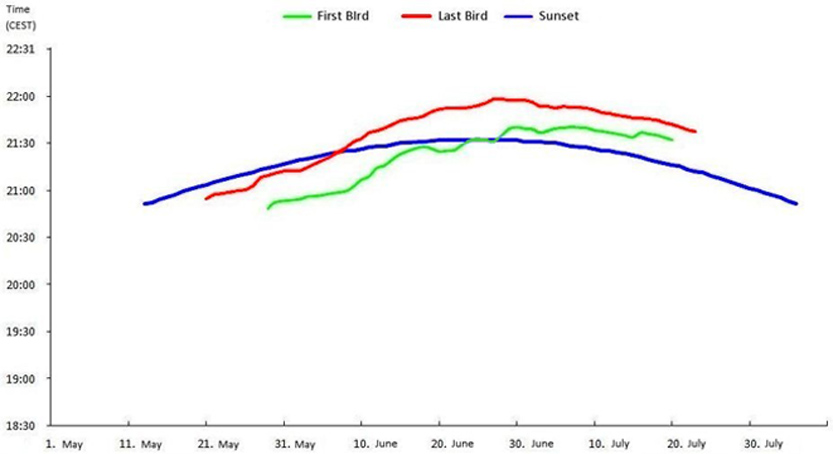
Figure 1. Times for the final evening return of breeding pairs, giving the time of sunset and light-intensity (Lux) for the second bird, with some weather and breeding data (W wind, R rain) 4. Discussion If one postulates the ideal situation of last entering, e.g. under warm weather conditions and a cloudless sky, one would obtain a smooth graphical curve. Another possibility to obtain the sense of a behaviour is by smoothing out the data. This is portrayed in Figure 2. In these curves the value of each day is the average of the 11 preceding and 11 following days. With this method one can clearly see the tendency to a later return with the progress of the growth of the young. In reality of course there are deviations (Figure 1) which can be explained by the following factors.
Figure 2. Smoothed data showing changes in the time of last return to nest. The flight activity of breeders increases with dawn as hatching progresses (Legend see table 1; smoothing factor K 23) 4.1. Light Intensity When the weather is clear and bright, with the exception of the following cases, the return of the second bird takes place exclusively at or immediately after sunset. From 155 returns to nest observed, 40 took place before and 115 after sunset. So sunset is not a fixed quantity for the birds. The times of flying activity are adapted rather to prevailing conditions. Before the eggs were laid the breeding pair flew in within a range of light intensity between 77 and 10900 Lux, with a tendency towards values between 77 and 176 Lux, during the period of breeding values between 17 and 360 Lux, with a greater tendency to lower values between 17 and 73 Lux. During the feeding period values between 8 and 159 Lux were measured, the majority between 17 and 20 Lux (35 % ). On the days of flight of the fledglings neither parent returned for the night. After the fledglings had achieved flight the values fell to between 9 and 18 Lux. 4.1.1. Weather factors The influence of the weather is clearly noticeable. In 14 cases rainy weather, rain or thunderstorms (all R in table 1) were the reason for an early return to the nest. In comparison with rainless days, the return was between 9 and 59 minutes (average 25 minutes, n=14) earlier and was in fact irrespective of the light intensity. One reason for the early return can be found in the fact that the density of airborne insects is low when the weather is rainy and a feeding flight for swifts is not suitable. This observation is quite interesting and suggest that social behavioural patterns are less important for late flying than the need to conserve energy when adverse insect availability conditions prevail. The influence of windy or squally weather was recorded only three times. On the evening of July 14th both swifts returned 67 and 54 minutes earlier in comparison with a cloudy July 17th, despite a 36 Lux higher light factor. On the two other days (24.06. and 22.07.) the early return time was diminished by about 10 minutes. 4.1.2. Clouding over The effects of weather, described in the preceding section, are caused, with the exception of the wind factor, by a clouding over and thus are linked to reduced light intensity. Early returns which were exclusively caused by clouds were registered only twice and occurred on 16.05. and 06.07. I failed to measure the Lux on May 16th because I did not expected the last return at the early hour of 20.34. 4.1.3. Social Factors For further nine deviations from the postulated ideal line there are no obvious physical causes. These took place on May 5th and 19th with fine weather (1050 res. 374 Lux), May 21st, 22nd and 23rd with blue sky, May 25v and 27th (cloudless, 10900 Lux, res. clouded and 4240 Lux) and July 2nd and 4th (each day cloudy, 36 and 20 Lux). The temperature range on those days lay between 8 and 19° C minimum, and between 18 and 28° C maximum and so were not unusual. With the exception of May 5th, which was the day of return from Africa, social factors offer possible explanations. On May 26th and 28th the eggs were laid, lending credence to an early return to nest the evening before. Naturally this observation of only two dates lies within the limits of statistical error. But it is interesting that this behavior was noted on the two evenings before egg laying and was exhibited by both partners. Moreover the return on May 25th took place at the same time. On the evening before the third egg was laid, the weather was poor so that this early return can not be taken as a sole reason for this behavior. But the same behavior can be noted in the graphics presented by Gory (1992). In five of the breeding pairs he observed that the first bird entered earlier on the evening before the eggs were laid. The following events also speak in favor of social factors playing a role in early return to nest. May 21st was day five before the laying of the first egg. This is the usual time for the gestation of swift eggs (Lack 1956, Weitnauer & Scherner in Glutz von Blotzheim 1994), so it can be assumed that mating occurred on the respective evenings. The same is true for May 19th and the following days. In Gory’s graphics one can find at least one early return on the fifth day before egg laying. Finally July 2nd and 4th were the evenings before the chicks opened their eyes. The parents reacted to this incident in both cases. Because this occurred during the period of permanent presence in the nest Gory’s methods of measuring could not record it. The behavior of entering is remarkably similar with both members of the pair. On ten days they entered the hole consecutively with a lapse of under a second. Sometimes I was able to watch them flying so close behind one another towards the hole that one bird had to break the flight just to let the partner enter. The distance in those cases must have been even closer than in previously described cased. The human eye cannot differentiate with such precision. On 13 days both partners returned within two minutes of each other from their last flight. The biggest synchronisation was noted in the period without brood duties, such as before breeding and again four days before the fledglings took first flight. Around this time the phase of feeding the young by the parents is concluded (Schulz 1881, Weitnauer 1947, Lack & Lack 1951, Wendt 1988, Polaschek 1992). In the period between the egg laying and the eleventh day after hatching both parents entered the nest box together only once. This is the phase of the nearly continuously brooding (Cramp 1994, Gory 1992). It can be stated that swifts demonstrate impressive social contact. In this context some answers to the question concerning a possible differentiated role played by the two sexes with relation to the last return to nest would be of great interest. While there are occasionally some statements about the bird’s sex in the literature (Weitnauer 1947, Zimmermann 1956, Flath 1980, Bezzel 1985), it is not made clear how the sexes are identified and were recognised. Such remarks are normally without value since live males and females can only be identified when copulating or , a recent discovery, when screaming (Kaiser 1997). To answer the above question the birds would have to be marked in a manner that which would make verification possible throughout the season without repeated catching, which is impossible under current protection laws. In the investigations presented here the sex of the birds was not determined. The swifts were numbered by the sequence of entering. The last home-comer to the nest might have been the male or the female. So far - apart from time when copulation occurs and the calling pattern - it is not yet known whether there is a difference in behavior in the male and female swift, particularly concerning the sequence of the last evening entering of the nest. Acknowledgments I wish to thank Dr. Henry Hahnke, Museum of Natural History, Berlin for valuable criticism and Prof. Brian Hawkins, Berlin for the linguistic revision of the English text. Note. Scheer 1949 published values of measurements of sun rising and morning flight in Darmstadt. The values are calculated again and are between 18-29 Lux under good and between 103-165 Lux under bad weather conditions. References
Auel, H. 1917: Beobachtungen über die Potsdamer Vogelfauna und über das Erscheinen ihrer Zugvögel im Frühjahr. Journal für Ornithologie 65:87-92 Bezzel, Einhard 1984: Kompendium der Vögel Mitteleuropas. Nonpasseriformes, p. 674. Wiesbaden Church, H.F. 1956: The Roosting Times of the Swift. Bird Study 3:217-220 Cramp, Stanley (Hrsg.) 1994: Handbook of the Birds of Europe, the Middle East and North Africa, Bd. 4. Oxford-New York, p. 657-670 Daanje, A. 1944: De Vliegende Gierzwaluw, Apus a. apus (L.). Ardea 33:74-84 Gory, Gérard 1992: Activité au nid du Martinet noir Apus apus (L.), durant la période de couvaison. Alauda 60:134-142 Guérin, G. 1923: La vitesse de vol des oiseaux et l’aviation. Revue francaise d’Ornithologie 8:74-79 Haartman, Lars von 1940: Über den Tagesrhythmus des Mauerseglers, Apus a. apus (L.). Ornis Fennica 17:7-11 Haartman, Lars von 1949: Neue Studien über den Tagesrhythmus des Mauersgelers, Apus apus (L.). Ornis Fennica 26:16-24 Hoffmann, B. 1917: Der Gesang und Andres von unserm Mauersegler (Cypselus apus L.). Journal für Ornithologie 65:459-464 Kaiser, Erich 1997: Sexual recognition of Common Swifts. British Birds 90:167-174 Keller, Dietmar 1977: Messung der Aktivität des Mauerseglers an der Bruthöhle. Der Falke 24:242-245 Kumerloeve, H. 1954: Segler als Nachtflieger. Die gefiederte Welt 78:98 Lack, David & Lack, Elizabeth 1951: The breeding biology of the swift Apus apus. Ibis 93:501-546 Lack, David & Lack, Elizabeth 1952: The breeding behaviour of the Swift. British Birds 45:186-215 Lack, David 1956: Further notes on the breeding biology of the swift Apus apus. Ibis 98:606-619 Litzbarski, H.: Mauersegler. In Rutschke, Erich (Hrsg.) 1987: Die Vogelwelt Brandenburgs. p. 252-253. Wiesbaden Polaschek, Ingeborg 1992: Elternlose Jungvögel. Niedernhausen/Ts., p. 51-52 Quantz, B[ernhard] 1917: Vom Zuge des Mauerseglers in Göttingen (Südhannover). Die Gefiederte Welt 46:197-199 Scheer, Georg 1949: Beobachtungen über den morgendlichen Flugbeginn des Mauerseglers, Micropus apus (L.). Die Vogelwarte 15:104-109 Schulz, E. 1881: Eine junge Turmschwalbe (Cypselus apus). Ornithologisches Centralblatt 6:126 Schuster, Ludwig 1921: Über Zuruhegehen und Abzug des Turmseglers im Sommer 1920. Ornithologische Monatsberichte 29:30-31 Sehlbach, Fr. 1926: Beobachtungen an Turmschwalben. Ornithologische Monatsschrift 51:40-42 Spallanzani, Lazzar 1797: Viaggi alle due Sicilie e in alcune parti degli Apenini 6. Pavia. Cit. after Weitnauer, Emil & Scherner, Erwin. R.: Apus apus. In Glutz von Blotzheim (Hrsg.)1994: Handbuch der Vögel Mitteleuropas, p. 671-712. Wiesbaden Swaen, A.E.H. 1922: Bijzonderheden betreffende de Gierzwaluw [Apus apus (L.)]. Ardea 11:117-123 Tigges, Ulrich 1994: Beobachtungen am Mauersegler (Apus apus) und Bericht über eine Spätbrut 1993. Berliner ornithologischer Bericht 4:129-141 Vleugel, D.A. 1947: De duur van de vliegdag van de Gierzwaluw, Apus a. apus (L.). Ardea 35:222-224 Wendt, Ernst 1988: Zum Ausfliegen junger Mauersegler (apus apus). Die Vogelwelt 109:128-129 Weitnauer, Emil 1947: Am Neste des Mauerseglers, Apus apus apus (L.). Der Ornithologische Beobachter 44:133-182 (Supplement) Weitnauer, Emil 1952: Uebernachtet der Mauersegler, Apus apus (L.) in der Luft? Der Ornithologische Beobachter 49:37-44 Weitnauer, Emil 1954: Weiterer Beitrag zur Frage des Nächtigens beim Mauersegler, Apus apus. Der Ornithologische Beobachter 51:66-71 Weitnauer, Emil 1960: Über die Nachtflüge des Mauerseglers, Apus apus. Der Ornithologische Beobachter 57:133-141 Weitnauer, Emil & Scherner, Erwin. R.: Apus apus. In Glutz von Blotzheim (Hrsg.)1994: Handbuch der Vögel Mitteleuropas, p. 671-712. Wiesbaden Zimmermann, Herbert 1956: Brutbiologisches vom Mauersegler (Micropus apus). Ornithologische Mitteilungen 8:89-90 |
|
Received
11.06.1997 - Accepted 05.12.2000
|
|
© APUSlife
2000, No. 2340
The German version “Das täglich letzte Einfliegen der Mauersegler (Apus apus) in Abhängigkeit von Brutphänologie, Umwelt- und sozialen Faktoren“ (APUSlist No. 0067) is to be published simultaneously in Berliner ornithologischer Bericht 09. |
![]()
![]()
![]()