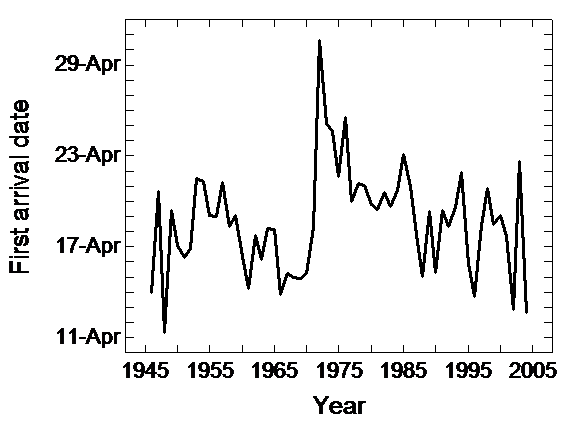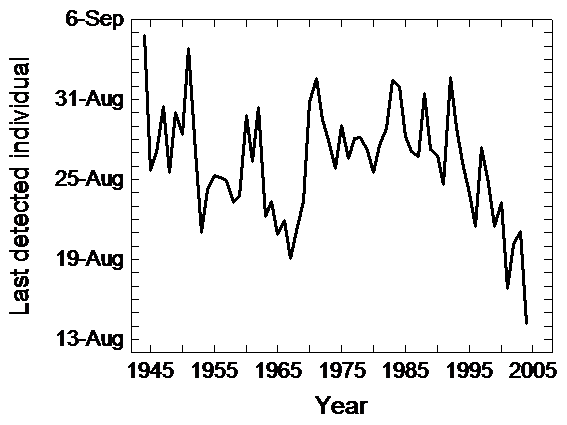|
Abstract Every year, millions of migratory birds visit Europe. Their arrival and departure dates ave now been recorded over many decades providing a valuable source of information to test whether present climate change is affecting migratory bird phenology. Among migrant birds, the Common Swift is one of the most widespread and abundant species in the Western Palearctic, which has favoured the existence of many long term studies about its migratory phenology. A review of literature yielded 16 studies that have reported time-series of Common Swift migratory phenology. A significant advancement of the arrival dates was reported only in 6 of them. Arrival dates were related to climate only in 4 out of 16 studies and thus the attribution of such advancements to climate change is difficult. In some studies, the departure date is advancing and the length of stay increasing, but other studies do not confirm this. Only 5 studies have assessed temporal trends both for departures and length of stays and consequently results for both variables are far from conclusive. No definitive answer to the question of whether climate change has affected the Common Swift migratory phenology can be given. |
|
OSCAR GORDO: |
||||||||||||||||||||||||||||||||||||||||||||||||
|
Does climate change affect Common Swift Apus apus migratory phenology? |
||||||||||||||||||||||||||||||||||||||||||||||||
|
Introduction: impacts of climate change on birds
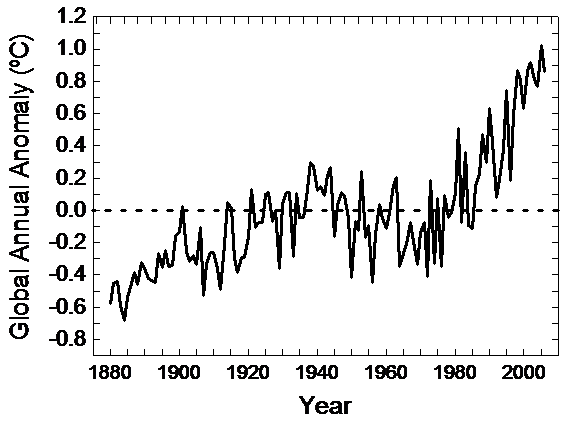
The Earth’s climate is changing (IPCC, 2001). This change is often reported as an increase of temperatures, so called global warming, resulting from the increase in atmospheric concentration of greenhouse gases due to human activities (Crowley, 2000). During the last century global temperatures have increased on average about 0.6 ºC (Fig. 1). However, alterations in other climatic features, such as rainfall (Milly et al., 2005), extreme weather events (Easterling et al., 2000), or global atmospheric circulatory systems (Hurrell, 1995) are also expected.
The alteration of climatic patterns in recent decades has already caused consequences for organisms (Hughes, 2000; McCarty, 2001; Peñuelas & Filella, 2001; IPCC, 2002; Walther et al., 2002; Parmesan & Yohe, 2003; Root et al., 2003; Root et al., 2005). One of the most evident of such changes has been the alteration of seasonal timings in northern latitudes. At present, spring is coming earlier while autumn is delaying its arrival (e.g., Menzel & Fabian, 1999). The potential effects of these changes on organisms’ life-cycles are large and range from ecosystem functioning to species competitive abilities.
In the case of birds, many studies have demonstrated long-term shifts in many biological parameters (see Table 1), in most cases attributed to the recent climate change (Crick, 2004). In fact, the effect of climate on most of these parameters has been well-known for many decades, but this knowledge has become extremely useful recently for the assessment and prediction of climate change impacts. However, some other environmental changes acting on a long-term temporal scale (e.g., land-use changes, habitat fragmentation) should be also taken into account as possible drivers of such changes (or absence of them).
Species’ distribution, both in latitude and altitude, is strongly constrained by climate. Therefore, it is expected that birds will react to climatic changes by finding new suitable places according to their ecological requirements. Thomas & Lennon (1999) reported an average northward movement of 19 km of the northern margin of British breeding birds in just 20 years, between 1968-72 and 1988-91. They suggested that this northward expansion is probably due to the climatic warming experienced during the study period since temperatures are closely related to the breeding success, distribution and diversity of British birds.
Abundance of birds is strongly determined by environmental conditions and climate is one of the most important regulatory non-biological mechanisms to determine the number of individuals and their potential increase (Stenseth et al. 2002). Since each species is affected by different key climatic variables, the same change can benefit one but harm another. Sæther et al. (2000) demonstrated that an increase of winter temperatures strongly affected the dynamics of a dipper (Cinclus cinclus) population from southern Norway. Low recruitment and population size decrease was found after cold winters which were associated with a higher number of days with ice cover in the study area which impaired foraging opportunities for this species. In Scotland, the number of capercaillies (Tetrao urogallus) has been decreasing for three decades. Moss et al. (2001) examined whether climate change was a factor in this population decline; interestingly, temperatures have fallen during a critical period in spring for chick survival in the study area. Since reproductive success is dependent on spring weather, the number of reared chicks per female has decreased from 3 to close to 0 and population viability is therefore endangered.
Climatic changes also can have impacts on individuals’ morphology. Yom-Tov (2001) studied the specimens of resident passerines from the Zoological Museum of the Tel-Aviv University collected during the second half of the 20th century in Israel. He found declines in body mass in four out of five species studied (20% in the graceful warbler Prinia gracilis, 9.7% in the house sparrow Passer domesticus, 18.6% in the yellow-vented bulbul Pycnonotus xanthopygos and 20.8% in the Sardinian warbler Sylvia melanocephala). He suggested that such declines were probably a response to an increase of 1.3 ºC in temperatures in Israel during the second half of the 20th century reflecting Bergmann’s rule (i.e. individuals from warm regions are smaller than those from cold regions in warm-blooded animals). Møller & Szép (2005) studied the increase in the length of the outermost tail feathers of barn swallow Hirundo rustica males from a Danish population from 1984 to 2003. This secondary sexual character changed in response to the deteriorating ecological conditions in northern Africa, an important staging area during spring migration for the studied population.
The timing of life-cycle events is one of the most obvious shifts due to climate change. Crick et al. (1997) showed that 51 out of 65 species of U.K. birds tended to lay eggs earlier during the period 1971-95. In these 51 species, 20 showed significant advances that ranged from 4 to 17 days. They suggested that the observed change was a response to the increase of spring temperature and the advancement of food availability (e.g., arthropods). Migratory phenology has also shifted during the last decades (e.g., Sparks, 1999). In the particular case of migratory species, the advancement of spring in their breeding grounds (e.g., Menzel & Fabian, 1999) has been proposed as a reason for the observed advancement in their arrival dates in recent decades. An early arrival has benefits for individuals in terms of better territories and mates, or increased offspring survival (Møller, 1994). It is not surprising that migratory birds are reacting to climate change to profit from the opportunities offered by a phenologically advanced scenario. However, the timing of life-cycle events of migratory birds is also controlled by endogenous rhythms (Berthold, 1996) which ensure that individuals perform each phase of their life-cycle (e.g., migration, reproduction, moult) at the right time. Such endogenous rhythms are in turn synchronized with the annual seasonality through the most reliable calendar: the photoperiod. Since endogenous rhythms are not controlled by ambient temperature, migratory species could be constrained in providing an adequate response to climate change challenges (Both & Visser, 2001).
The Common Swift Apus apus is one of the most abundant and widespread trans-Saharan bird species in the western Palaearctic. Hence, it is not surprising that the Common Swift has received much attention since many decades (e.g., Lack, 1956) and, consequently, there are many long-term time-series about migratory phenology of this species that have allowed us to determine the potential effects of recent climate change on Common Swift. Here, I review these results obtained for this species in several published studies to date but it must be noted that in most cases these studies were not focussed solely on this species and consequently appropriate interpretation of the results obtained for the Common Swift is lacking. In the same way, the heterogeneity of the type and temporal samples of such studies can also be a handicap for perfect comparison. However, in browsing together all the published results, I seek to offer an overview of the long-term changes reported as affecting this species.
Studies in the United Kingdom
Many studies have analysed long-term changes of bird migratory phenology in the U.K. based on county and observatory reports. Mason (1995) analysed the first arrival date of 23 migrant bird species to Leicestershire for the period 1942-91. He showed that the arrival dates of the Common Swift were strongly fitted to the last week of April, i.e. showed low variability year to year. The arrival of Common Swifts to Leicestershire did not change during the study period, although a slight advancement was detected during the last two decades studied. The spring arrival of migrants (measured as the first day on which a species was recorded) to Leicestershire were reassessed together with records from Sussex by Loxton et al. (1998) between 1966 and 1996. As in the previous study, no temporal trend was found for the arrival date of the Common Swift to Leicestershire, nor to Sussex. In both areas, the arrival date of Common Swifts was not related to spring Central England temperature, which is broadly applicable to the U.K. Loxton et al. (1998) found only a significant increase of Central England temperatures for April (r = 0.352) and March-April (r = 0.442) during the period 1966-1996. Loxton & Sparks (1999) examined the first arrival dates of 24 species at four bird observatories on the western coast of England and Wales (Portland, Skokholm, Bardsey, and Calf of Man) during the period 1946-98. Only arrivals at Portland since 1969 showed a significant advancement (change rate not reported). No significant relationship was found with Central England temperatures at February, March or April in any case. Interestingly, a recent reassessment of Portland records for the period 1959-2005 (Croxton et al., 2006) showed a non-significant trend of -0.77 days·decade-1, while it was a bit steeper for the period 1982-2005 with a trend of -0.99 days·decade-1 (n.s.). In this new study, authors found that first arrivals were more closely related to April temperatures in Spain (r = -0.49, p < 0.05) and France (r = -0.46, p < 0.05) than to Central England temperatures (r = -0.24, n.s.). First Swifts arrive at Portland 3.22 days earlier per each increased degree in April temperatures from Spain.
Jenkins & Watson (2000) analysed the first arrival date or the first song heard in 35 species, both of migrants and residents, during the period 1974-99 near Aboyne village (50 km west of Aberdeen). They reported an earlier but non-significant trend toward earlier arrivals for the Common Swift (Spearman’s correlation = -0.28; n= 18). Sparks & Mason (2001) studied arrival and departure (measured as the first and last detected migrants) dates of 39 migratory birds at Essex for the period 1950-98. Both arrivals and departures of Common Swifts changed significantly over the last decades. An advancement of 2 days per decade for arrivals and a delay of 5 days per decade for departures, enlarged the length of stay in the U.K. at a rate of about 6 days per decade. Surprisingly, temporal trends in arrival and departure dates were not related to temperatures during the spring nor the summer respectively, which makes difficult the attribution of such phenological shifts to climate change.
Cotton (2003) studied the arrival and departure dates, measured as the first and last individual, of 20 migratory birds at Oxfordshire during the period 1971-2000. In the case of the Common Swift, he reported advancement both in arrival and departure dates (3 days·decade-1 and 0.6 days·decade-1, respectively). This result concurs with those reported by Sparks & Mason (2001) for arrivals but is the opposite for departures. Consequently, Cotton (2003) reported a lesser increase in the length of stay (2 days·decade-1). The different study period and area could account for these differences. However, the latter author concluded that the timing of arrival had advanced (1.53 days·ºC-1) in relation to increasing winter temperatures in sub-Saharan Africa, whereas departure dates had advanced (-0.06 days·ºC-1) in response to increased temperatures in Oxfordshire during the 30 year study period.
Summarising, most of the analysed time-series for spring arrival dates showed non-significant temporal trends, while the two reported trends for the autumn departures were opposite. There is little evidence for changes in the duration of stay in the U.K but it could be increasing.
Studies in Central and Northern Europe
On the German island of Heligoland (North Sea), a monitoring programme of bird migration based on standardized trapping and counts was started in 1961 (Bairlein & Winkel, 2001). Twenty-one regular bird species on passage on the island showed considerable changes in the timing of migration. In the particular case of the Common Swift, an advancement both in median dates of passage during spring and autumn was recorded during the study period (-0.35 days·decade-1 and -0.88 days·decade-1, respectively), although only the advancement of autumn passage was strongly significant (p<0.001). The time span between the mean spring and autumn arrivals was reduced (-0.85 days·decade-1). In the same country, Witt (2004) studied the first and last records of the Common Swift in Berlin for the period 1965-2002. No significant trend was found in any phase (r=-0.21 for arrivals, r=0.07 for departures). The author concluded that the effects of climate change were not evident, in spite of the increase of annual mean temperatures in Germany of about 1.4 ºC for the period 1961-2000 (Chmielewski et al., 2004).
In Lithuania, Zalakevicius et al. (2006) examined the first arrival date of 40 species in the environs of Vilnius over a period from 1971 to 2004 in daily fixed 6 hours duration transects. In the particular case of the Common Swift, a significant trend toward earlier arrivals was detected during the study period (-3.7 days·decade-1). Interannual fluctuations in arrivals were negatively related to April temperatures at Vilnius and at Prague (Czech Republic) but not to precipitation, atmospheric pressure or the North Atlantic Oscillation (NAO) weather system. Air temperatures for the eastern Baltic region rose by 0.45 ºC during the last century. This increase was especially sharp for winter and spring (up to 1.7-2.0 ºC warmer conditions). The absence of significant relationship between Common Swift arrivals and NAO had been previously reported for long-term records from South Moravia (Czech Republic; Hubálek, 2003), and from the Finnish observatories of Jurmo and Hanko (Vähätalo et al., 2004).
In their review, Lehikoinen and colleagues (2004) studied the arrival date of six common migrants recorded at Turku (Finland) since 1749. The impressive time-series for the Common Swift (the longest in the world, to the best of my knowledge) shows no clear trend (not reported by authors) during the last decades. The authors demonstrated that very long time-series often show periods of constant direction of change (i.e. advance or delay) in arrival dates. Therefore, historical records offer an important lesson: obtained trends from short time-series could be merely fluctuations around the overall average arrival dates of a certain species.
In summary, central and northern European populations of Common Swifts did not show clear trends in arrival dates or in autumn departures, in recent decades. Most trends for arrivals were negative (i.e. advancement) but only in Lithuania did such changes achieve statistical significance.
Studies in Southern Europe
In the Mediterranean basin, the first evidences about long-term changes in bird migratory phenology were reported by Peñuelas et al. (2002). These authors analysed the first arrival date of six common trans-Saharan migrant bird species at one locality in north-eastern Spain for the period 1952-2000. Interestingly, all species except the Common Swift tended to arrive later. This dataset was reassessed by Gordo et al. (2005) in order to examine the reason for such temporal trends in the light of the overwhelming evidence of advancement in spring migration timing. Moreover, annual temperature in the study site during the study period increased about 1.5 ºC, and plant and insect phenology advanced their spring events (leaf unfolding -3.2 days·decade-1, flowering -1.2 days·decade-1 or insect appearance -2.2 days·decade-1). The study of Gordo et al. (2005) showed that arrivals of migrant birds were always related to climatic patterns in Africa, but never to climate in the study locality. Rainy and warm years in Africa advanced the first detection of migrant birds in the locality. In the particular case of the Common Swift, precipitation patterns in Eastern Africa were the most closely related meteorological variable to the recorded arrivals in north-eastern Spain, however their effect was very low (r2 = 0.087, p = 0.048).
In another study, Gordo & Sanz (2005) analysed long-term changes in plant and animal phenology for the Ebro Observatory (Tortosa, north-eastern Spain). An increase of about 1 ºC of annual mean temperatures during the last 50 years at this site was recorded. In opposition to the previous findings, 3 out of 4 trans-Saharan bird species advanced their arrivals during the last three decades. In the case of the Common Swift, first sightings shifted towards earlier dates and last sightings toward later ones. As result, the total number of days between first and last sighted individuals has increased since the mid-1970s. Recently, Rubolini et al. (2007) found similar results for a breeding colony of ca. 30 pairs from central-eastern Lombardy (Italy). First arriving individuals advanced their dates for the period 1982-2006 at a rate of -3 days·decade-1, and their arrivals were strongly related to April temperatures at the breeding site (-3.84 days·ºC-1). The authors suggest that Common Swifts are reacting to increasing April temperatures (1.25 ºC during the study period) by progressively advancing their arrival dates.
Another study by Gordo & Sanz (2006) analysed first arrivals, last departures and length of stays of five common trans-Saharan bird species for the whole of Spain for the period 1944-2004. Since this study relied on thousands of Spanish localities, it could offer a more conclusive picture of temporal changes in migratory phenology in the Mediterranean region than previously mentioned studies. Local studies are useful because in most cases they are the only available data but peculiarities related to each local population could explain the idiosyncrasy found in phenological responses. For the whole of Spain, Gordo & Sanz study showed an advance in the arrival date of the Common Swift since the 1970s (Fig. 2). However, more recent arrival dates are quite similar to those reported prior to the 1960s. The study, therefore, concluded that the advancement recorded during the last three decades should be interpreted as a re-establishment of the normal pattern of migration after an anomalous period of delayed arrivals, especially during the 1970s. Spring phenology was significantly related both to climatic fluctuations in Spain and Africa. The higher the temperatures in spring in Spain the earlier birds arrived there. In the same way, rainy winters in western Africa were also associated with earlier arrivals to Spain in the next spring. The enormous delay in arrivals recorded in 1972 was due to the combination of extreme climatic conditions in the breeding and wintering grounds simultaneously, with very low temperatures in Spain and very scarce precipitation in western Africa. On the other hand, last sighting dates and the total number of days between the sighting of the first and last individual of each year did not show significant trends for the overall period. Nevertheless, since the mid-1980s the last individuals sighted have been recorded earlier (Fig. 3) and the length of stay as shorter. However, interannual fluctuations in the last sightings of Swifts were not significantly related to any climatic parameter, nor temperature at departure time nor temperature during reproduction. Due to the southern location of the Iberian Peninsula within Europe, some of the last reported individuals could be passage individuals from northern populations and this may hamper an accurate assessment of autumn migration in Spain on basis to field observations of volunteers.
In summary, there has been an advancement in arrival dates in the last decades but taking the longer view arrival dates appear to be returning towards "normal" migratory dates after an anomalous period of very late arrivals in the 1970s. Departure dates did not show changes for Spain as a whole, although at a local level advancement has been recorded.
Conclusions
There are no clear temporal trends in Common Swift migratory phenology in the 16 studies which have reported results for this species. First arrival dates did not show significant changes during last decades in most cases. However, those studies reporting a significant trend (Sparks & Mason, 2001; Cotton, 2003; Zalakevicius et al., 2006; Gordo & Sanz, 2005; Gordo & Sanz, 2006; Rubolini et al., 2007) concur in recording earlier dates in recent years. This result would be in agreement with a hypothesis based on an advancement of spring migration in response to climate change. However, in a few studies, these observed temporal trends in arrival dates were significantly related to climate variables (Cotton, 2003; Zalakevicius et al., 2006; Gordo & Sanz, 2006; Rubolini et al., 2007), and thus the attribution to climate change is difficult. Temporal trends in departure dates were reported in far fewer cases (only in five studies), but in two out of five cases, significantly earlier dates were recorded in timing of the birds’ departure from the breeding grounds. Since the Common Swift produces only one brood per year, this advancement in post-breeding migration could be due to a general advancement of the whole breeding period of the species (Ellegren, 1990). Despite the parallel trend towards an advancement both in arrivals and departures, significant results for the number of days that the species is observable in their breeding grounds indicated an increase in the length of stay (Sparks & Mason, 2001; Cotton, 2003). It could be concluded that arrivals are moving towards slightly earlier dates. The picture is even less conclusive for both departures and stays but few studies have analysed long-term changes in departure dates and lengths of stay for this species. The idiosyncrasy of reported results may be due to the heterogeneity of reviewed studies; the study years, size of geographical coverage, localisation and definition of phenological phases, varied across studies. For example, some studies analysing the total distribution of arrival dates in migratory birds have shown that temporal trends are different between earlier and later individuals of the population (Rainio et al., 2006; Tøttrup et al., 2006). In spite of a common trend towards earlier arrival dates for both first individuals and the bulk of the population, the arrival date of first individuals is even more advanced than that for the whole population. Consequently, those studies based on records of the first sighted individual could be overestimating the real advance in arrival dates of migratory birds. On the other hand, reported rates of phenological shift are strongly influenced by the choice of first and last years of studied time-series (Schaber & Badeck, 2005). Those time-series spanning the 1970s to the present show higher advancements than those spanning a wider temporal period (Lehikoinen et al., 2004) because the largest increase of temperature is indeed during the last three decades (Fig. 1). In conclusion, the diversity of studies is a serious handicap to comparison of results (Lehikoinen et al., 2004; Møller & Merilä, 2004; Tigges 2007) and there is no definitive answer to the question of whether climate change has affected the Common Swift migratory phenology.
I am very grateful to Mandy Mayer for polishing the English of this article.
References Archaux, Frederic (2004): Breeding upwards when climate is becoming warmer: no bird response in the French Alps. Ibis 146:138-144
Bairlein, Franz; Winkel, Wolfgang (2001): Birds and climate change. In: Lozan J.L., Graßl H, and Hupfer P. (ed.), Climate of the 21st Century: Changes and Risks. Scientific Facts, GEO, pp. 278-282
Berthold, Peter (1996): Control of bird migration. Chapman & Hall, London
Both, Christiaan; Visser, Marcel E. (2001): Adjustment to climate change is constrained by arrival date in a long-distance migrant bird. Nature 411:296-298
Brommer, Jon E. (2004): The range margins of northern birds shift polewards. Annales Zoologici Fennici 41:391-397
Chmielewski, Frank M.; Müller, Antje; Bruns, Ekko (2004): Climate changes and trends in phenology of fruit trees and field crops in Germany, 1961-2000. Agricultural and Forest Meteorology 121:69-78
Cotton, Peter A. (2003): Avian migration phenology and global climate change. Proceedings of the National Academy of Sciences USA 100:12219-12222
Crick, Humphrey Q.P.; Dudley, Caroline; Glue, David E.; Thomson, David L. (1997): UK birds are laying eggs earlier. Nature 388:527-527
Crick, Humphrey Q.P. (2004): The impact of climate change on birds. Ibis 146:48-56. Crowley, Thomas J. 2000. Causes of climate change over the past 1000 years. Science 289:270-277
Croxton, Philip J.; Sparks, Tim H.; Cade, Martin; Loxton, Richard G. (2006): Trends and temperature effects in the arrival of spring migrants in Portland (United Kingdom) 1959-2005. Acta Ornithologica 41:103-111
Dunn, Peter O. (2004): Breeding dates and reproductive performance. Advances in Ecological Research 35:69-87
Easterling, David R.; Meehl, Gerald A.; Parmesan, Camille; Changnon, Stanley A.; Karl, Thomas R.; Mearns, Linda O. (2000): Climate extremes: observations, modeling, and impacts. Nature 289:2068-2074
Ellegren, Hans (1990): Timing of autumn migration in bluethroats Luscinia svecica svecica depends on timing of breeding. Ornis Fennica 67:13-17
Gordo, Oscar; Sanz, Juan José (2005): Phenology and climate change: a long-term study in a Mediterranean locality. Oecologia 146:484-495
Gordo, Oscar; Sanz, Juan José (2006): Climate change and bird phenology: a long-term study in the Iberian Peninsula. Global Change Biology 12:1993-2004
Gordo, Oscar; Brotons, Lluís; Ferrer, Xavier; Comas, Pere (2005): Do changes in climate patterns in wintering areas affect the timing of the spring arrival of trans-Saharan migrant birds? Global Change Biology 11:12-21
Hubálek, Zdeněk. (2003): Spring migration of birds in relation to North Atlantic Oscillation. Folia Zoologica 52:287-298
Hughes, Lesley (2000): Biological consequences of global warming: is the signal already. Trends in Ecology and Evolution 15:56-61
Huntley, Brian; Collingham, Yvonne C.; Green, Rhys E.; Hilton, Geoffrey M.; Rahbek, Carsten; Willis, Stephen G. (2006): Potential impacts of climatic change upon geographical distributions of birds. Ibis 148:8-28
Hurrell, James W. (1995): Decadal trends in the North Atlantic Oscillation: regional temperatures and precipitation. Science 269:676-679
IPCC (2001): Climate change 2001: impacts, adaptation, and vulnerability. In: McCarthy JJ, Canziani OF, Leary NA, Dokken DJ, White KS (eds), Contribution of Working Group II to the Third Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge
IPCC (2002): Climate change and biodiversity. In: Gitay H, Suárez A, Watson RT, Dokken DJ (eds), Technical Paper V of the Intergovernmental Panel on Climate Change. IPCC, Geneva
Jenkins, David; Watson, Adam (2000) Dates of first arrival and song of birds during 1974-99 in mid-Deeside, Scotland. Bird Study 47:249-251
Lack, David (1956): Swifts in a tower. Methuen & Co, London
Lehikoinen, Esa; Sparks, Tim H.; Zalakevicius, Mecislovas (2004): Arrival and departure dates. Advances in Ecological Research 35:1-31
Lemoine, Nicole; Böhning-Gaese, Katrin (2003): Potential impact of global climate changes on species richness of long-distance migrants. Conservation Biology 17:577-586
Loxton, Richard G.; Sparks, Tim H. (1999): Arrival of spring migrants at Portland, Skokholm, Bardsey and Calf of Man. Bardsey Observatory Report 42:105-142
Loxton, Richard G.; Sparks, Tim H.; Newnham JA. (1998): Spring arrivals dates of migrants in Sussex and Leicestershire (1966-1996). The Sussex Bird Report 50:181-196
Mason, Chris F. (1995): Long-term trends in the arrival dates of spring migrants. Bird Study 42:182-189
McCarty, John P. (2001): Ecological consequences of recent climate change. Conservation Biology 15:320-331
Menzel, Annette; Fabian, Peter (1999): Growing season extended in Europe. Nature 397:659-659
Milly, P.C.B.; Dunne, K.A.; Vecchia, A.V. (2005): Global pattern of trends in streamflow and water availability in a changing climate. Nature 438:347-350
Møller, Anders Pape (1994): Phenotype-dependent arrival time and its consequences in a migratory bird. Behavioural Ecology and Sociobiology 35:115-122
Møller, Anders Pape (2004): Protandry, sexual selection and climate change. Global Change Biology 10:2028-2035
Møller, Anders Pape; Merilä, Juha (2004): Analysis and interpretation of long-term studies investigating responses to climate change. Advances in Ecological Research 35:111-130
Møller, Anders Pape; Szép, Tibor (2005): Rapid evolutionary change in a secondary sexual character linked to climatic change. Journal of Evolutionary Biology 18:481-495
Moss, Robert; Oswald, James; Baines, David (2001): Climate change and breeding success: decline of the capercaillie in Scotland. Journal of Animal Ecology 70:47-61
Parmesan, Camille; Yohe, Gary (2003): A globally coherent fingerprint of climate change impacts across natural systems. Nature 421:37-42
Peñuelas, Josep; Filella, Iolanda (2001): Responses to a warming world. Science 294:793-794
Peñuelas, Josep; Filella, Iolanda; Comas, Pere (2002): Changed plant and animal life cycles from 1952 to 2000 in the Mediterranean region. Global Change Biology 8:531-544
Root, Terry L.; MacMynowski, Dena P.; Mastrandrea, Michael D.; Schneider, Stephen H. (2005): Human-modified temperatures induce species changes: Joint attribution. Proceedings of the National Academy of Sciences USA 102:7465-7469
Root, Terry L.; Price, Jeff T.; Hall, Kimberly R.; Schneider, Stephen H.; Rosenzweig Cynthia; Pounds, J. Alan (2003): Fingerprints of global warming on wild animals and plants. Nature 421:57-60
Rubolini, Diego; Ambrosini, Roberto; Caffi, Mario; Brichetti, Pierandrea; Armiraglio, Stefano; Saino, Nicola (2007): Long-term trends in first arrival and first egg laying dates of some migrant and resident bird species in northern Italy. International Journal of Biometeorology in press
Sæther, Bernt-Erik; Tufto, Jarle; Engen, Steinar; Jerstad, Kurt; Røstad, Ole Wiggo; Skåtan JE. (2000): Population dynamical consequences of climate change for a small temperate songbird. Science 287:855-856
Schaber, Jörg; Badeck, Franz-W. (2005): Plant phenology in Germany over the 20th century. Regional and Environmental Change 5:37-46
Sparks, Tim H. (1999): Phenology and the changing pattern of bird migration in Britain. International Journal of Biometeorology 42:134-138
Sparks, Tim H.; Mason, Chris F. (2001): Dates of arrivals and departures of spring migrants taken from Essex Bird Reports 1950-1998. Essex Bird Report, 1999:154-164
Stenseth, Nils Christian; Mysterud, Atle; Ottersen, Geir; Hurrell, James W.; Chan, Kung-Sik; Lima, Mauricio (2002): Ecological effects of climate fluctuations. Science 297:1292-1296
Thomas, Chris D.; Lennon, Jack J. (1999): Birds extend their ranges northwards. Nature 399:213-213
Thorup, Kasper; Tøttrup, Anders P.; Rahbek, Carsten (2007): Patterns of phenological changes in migratory birds. Oecologia in press
Tigges, Ulrich (2007): On the phenology of Common Swift Apus apus in Eurasia and the problem of the duration of stay. Podoces 2 in press
Tøttrup, Anders P.; Thorup, Kasper; Rahbek, Carsten (2006): Patterns of change in timing of spring migration in North European songbird populations. – Journal of Avian Biology 37:84-92
Tryjanowski, Piotr; Sparks, Tim H.; Kuczyński, L.; Kuźniak, Stanisław (2004): Should avian egg size increase as a result of global warming? A case study using the red-backed shrike (Lanius collurio). Journal of Ornithology 145:264-268
Tryjanowski, Piotr; Sparks, Tim H.; Profus, Piotr (2005): Uphill shifts in the distribution of the white stork Ciconia ciconia in southern Poland: the importance of nest quality. Diversity & Distributions 11:219-223
Vähätalo, Anssi V.; Rainio, Kalle; Lehikoinen, Aleksi; Lehikoinen, Esa (2004): Spring arrival of birds depends on the North Atlantic Oscillation. Journal of Avian Biology 35:210-216
Walther, Gian-Reto; Post, Eric; Convey, Peter; Menzel, Annette; Parmesan, Camille; Beebee, Trevor J.C.; Fromentin, Jean Marc; Hoegh-Guidberg, Ove; Bairlein, Franz (2002): Ecological response to recent climate change. Nature 416:389-395
Witt, Klaus (2004): Erst-und Letztbeobachtungen des Mauerseglers (Apus apus) in Berlin. Berliner Ornithologischer Bericht 14:186-192
Yom-Tov, Yoram (2001): Global warming and body mass decline in Israeli passerine birds. Proceedings of the Royal Society of London-Series B 268:947-952
Zalakevicius, Mecislovas; Bartkeviciene, Galina; Raudonikis, Liutauras; Janulaitis, Justinas (2006): Spring arrival response to climate change in birds: a case study from eastern Europe. Journal of Ornithology 147:326-343 Table 1: Responses of several biological parameters of birds to climate change. A few example studies for such changes are offered. See reference list for the complete reference.
Figure 1
World annual temperature changes between 1880 and 2005. Solid line represents the difference of annual temperatures with the average temperature of the 1901-2000 period (dashed line). Negative values are colder years than 20th century average temperature, while positive values are warmer ones. Note the sharp trend toward warmer values since mid-1970s
Figure 2
First arrival date of Common Swift to Spain during the period 1944-2004. A trend toward earlier dates has been detected since mid-1970s. Nevertheless, arrival dates at present are similar to those reported previously to early 1970s.The strong delay recorded between 1970 and 1972 was due to the combined effect of weather conditions in Spain and Africa. Spanish springs were extremely cold and rainfall in the Sahel was extremely scarce Figure 3
Last detected individual date of Common Swift in Spain during the period 1944-2004. No temporal trend was detected during the six study decades. Nevertheless, departures are shifting toward earlier dates since mid-1980s. |
||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||
|
© APUSlife
2007, No. 3237 |
![]()
![]()
![]()