|
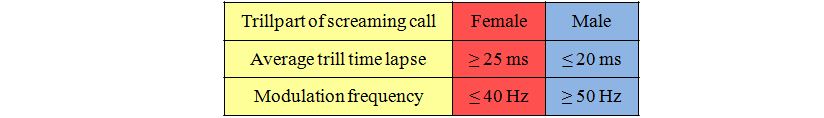
Abstract The typical screaming calls of the Common Swift (Apus apus) include a screaming part and a trill part. It will be shown that the pitch and the duration of the different parts of the screaming calls, produced by the single individuals of the breeding pairs in the nest box, vary intra- and inter-individually. The pitch and the length of the screaming calls are probably dependent on the degree of agitation of the Swifts and do not indicate the sex of the individual bird. The trill part codifies the sexual affiliation by the distances between the trills of a screaming call. Female Swifts show an average time lapse between the trills of ≥25 ms and males of ≤20 ms. This indicates that the fundamental frequency at the end of the screaming call of female Swifts is modulated by ≤40 Hz. Among male Swifts the modulation of the fundamental frequency is ≥50Hz. |
|
KONRAD ANSORGE:
|
| Sexual dimorphism of acoustic signals in the Common Swift Apus apus |
|
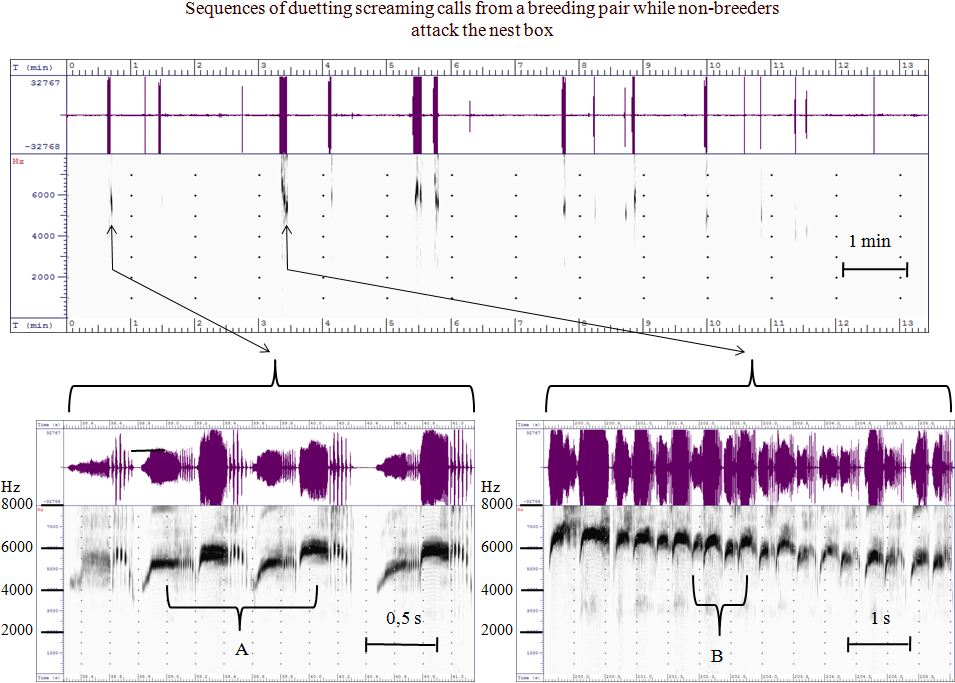
Introduction The Common Swift, Apus apus belongs to one of the best studied of bird species. There are, however, only a few publications which characterise and classify its vocal behaviour (Bretagnolle 1993, Malacarne et.al. 1989, van Oudheusden, 2006). Decades ago it had already been observed that the Common Swift typically demonstrates duetting. Lack (1956) described the partners of a pair alternately calling during the duet. One partner screams with a higher frequency than the other partner, this was named the swee-ree call. Based on the observations of Lack, Kaiser (1997) was able to conclude, by using a simple method of identification marks over several years of observation of his Swift colony, that the call of the female Swift sounds a little higher and the call of the male lower. He formed the opinion that he had found an acoustic signal which could enable the identification of the sex of a Common Swift. Bretagnolle (1993) also described an acoustic signal of sexual dimorphism. He stated that the screaming call always includes two parts: the screaming part and the trill part. He recognized that one partner of a pair has a faster trill rhythm, and the other one has a slower trill rhythm, and he hypothesised that this difference showed a sexual dimorphism. Furthermore, he assumed that male birds have the faster rhythm trill. Therefore Bretagnolle distinguished two types of screaming calls: type A (fast rhythm trill) and type B (slow rhythm trill). In the present investigation the results of Kaiser and Bretagnolle are examined by quantitative spectrographic methods. It is commonly observed that since that time the Swift population in Warsaw, as well as in other Polish cities and towns, has decreased considerably. Material and Methods In 2013 and 2014 three pairs of Common Swifts were observed in their nest boxes, where they were visible from the rear. Immediately after arriving each bird of a pair was marked with different colors on their wings and their screaming calls were recorded digitally and examined spectrographically. Based on this method the individual screaming calls were definitively characterised. The sexual affiliation of the single animals was determined during the egg deposition by film documentation to exclude mistakes. Either 10 single shots per second or film sequences with a length of 10 seconds were recorded. The sounds in the nest box were recorded by microphone capsules which had a frequency spectrum of 20 - 16000 Hz (KPCM-G60H50P-44DB-1185) and a digital recorder (Olympus LS-11). The sounds were analysed by different programs (SFS/WASP Version 1.54 und SFS/ESECTION of Mark Huckvale: University College London, Raven pro 64 1.4 und SRLab). A frequency of up to 8000 Hz was chosen for analysis and presentation in the spectrograms. The fundamental screaming frequency of the Common Swift is located within this frequency range. Further harmonics were omitted from analysis because they were unimportant for this investigation. Results and evaluation Analysis of the duetting Duetting means the alternate calling of a pair of Common Swifts. These sequences of screaming calls regularly occur at the nest when non-breeders pass the breeding box or even make contact with it while emitting loud screaming calls. This activity of non-breeders is called banging (Fig. 1). Such duetting appears to be employed to defend the breeding place (Bergmann 2014; Kaiser 1997). According to Bretagnolle (1993) duetting is individually distinctive and has a territorial function as well as being important for recognising an individual or the partner.
Fig. 2 shows two parts of the spectrogram of the vocalised duet sequences marked in Fig. 1. At the beginning of the banging (Fig. 2A) the male calls showed a 500 Hz lower frequency than the female.
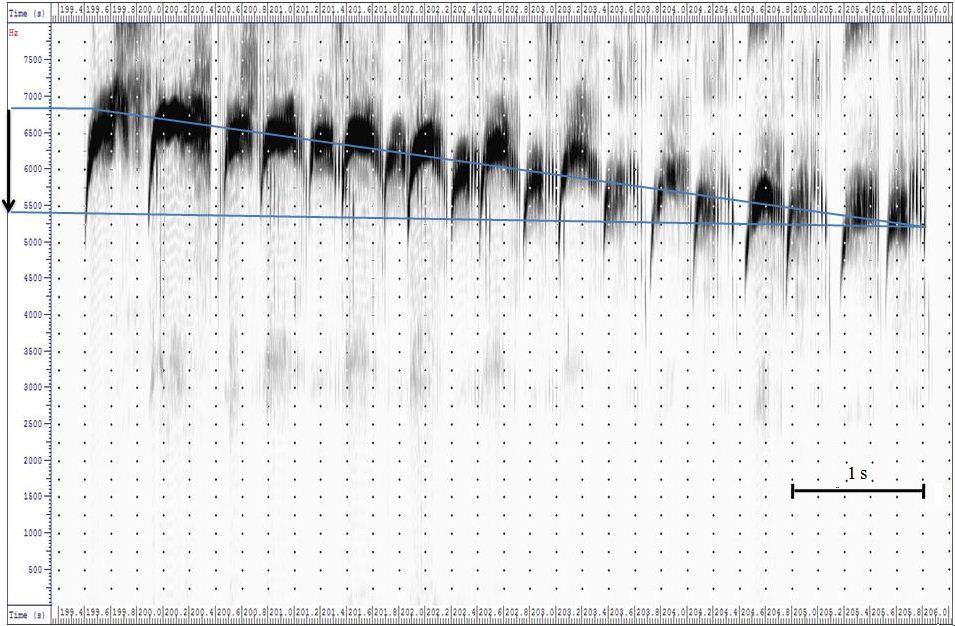
After repeated approaches from the outside, the intensity and duration of the duetting of the Swift pair in the nest box increases. Female and male Swift calls of higher frequency and intensity are emitted. Whereas the female only shows a slight increase in the frequency of 200Hz at the maximum, the male increases the frequency of the screaming call at about 1100 Hz and therefore exceeds the absolute pitch of the female by 400 Hz. (Fig. 2B) It is also remarkable that the total duration of the screaming calls of the two partners of the pair varies a lot. In the duets at the beginning of the "banging" the total duration of the screaming calls of the female is definitely longer (Fig. 2A) than in the duet, which is provoked 3 minutes later (Fig. 2B). In the course of the duetting the screaming calls of both partners assimilate more and more with the total duration and the frequency spectrum. The differences in the interval between the single trills during the trill part of the screaming calls remain the same Fig. 3 shows how the frequency of the screaming calls diminishes during the duetting. At the beginning the duetting starts at 6700Hz. At the end of the duetting, with reduced levels of agitation - the non-breeders have passed - the pitch of the screaming calls sinks to 5250 Hz.
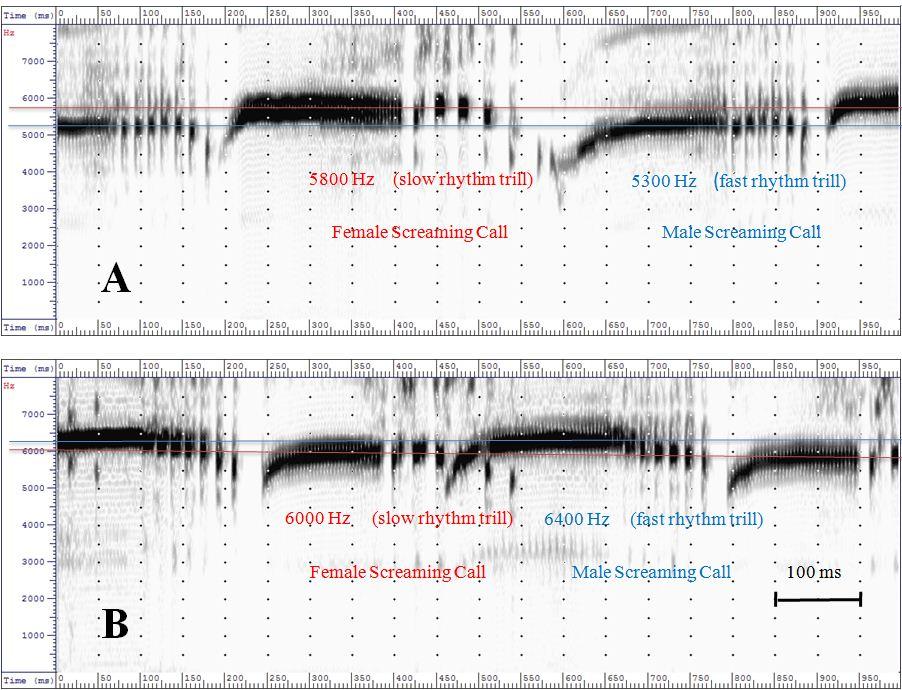
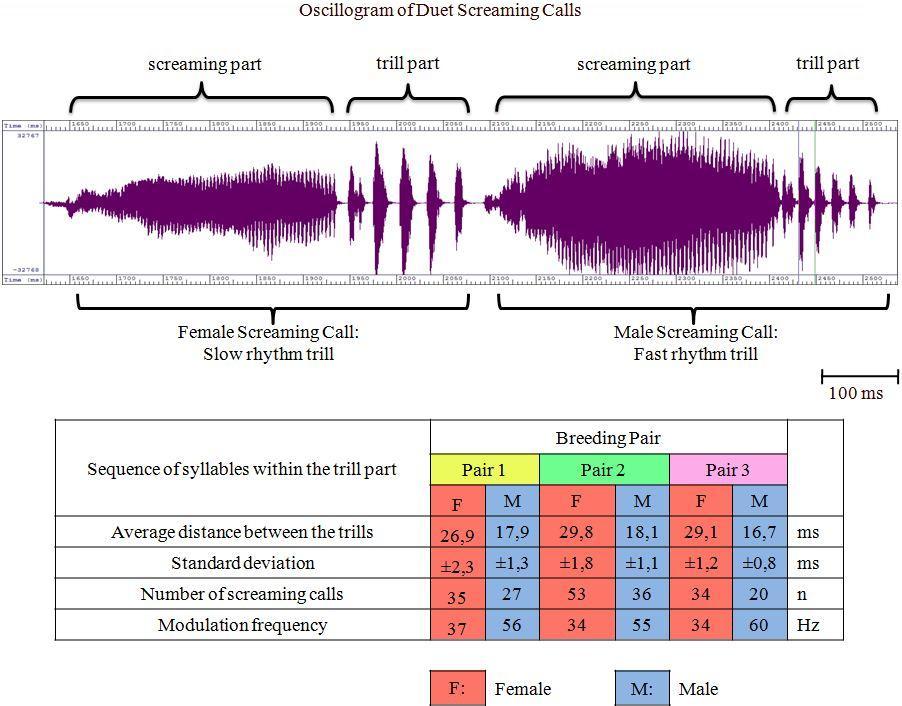
These observations were validated by observations of another pair of Swifts. In this case the male of the breeding pair even started the duet with a 250 Hz higher frequency than the female. In the course of the duet the frequencies assimilate more and more – as described before – and adapt with the pitch. The sex specific distinguishing feature of the calls is probably not codified by the frequency as described by Kaiser (1997). The variation in the duration of the screaming calls and their intensity probably reflects the agitation of the birds rather than providing evidence of sexual affiliation. Codifying of sex The codifying of the sexual affiliation seems to be identifiable within the trill part. The frequency of the screaming call – which serves as the carrier frequency – is modulated by a low frequency – the sequence of the trills. This indicates a frequency modulation akin to that used, for example, in ultra high frequency radio engineering. Measuring the temporal sequences of the screaming calls in the trill part of female and male Swifts leads to the following results (Fig. 4)
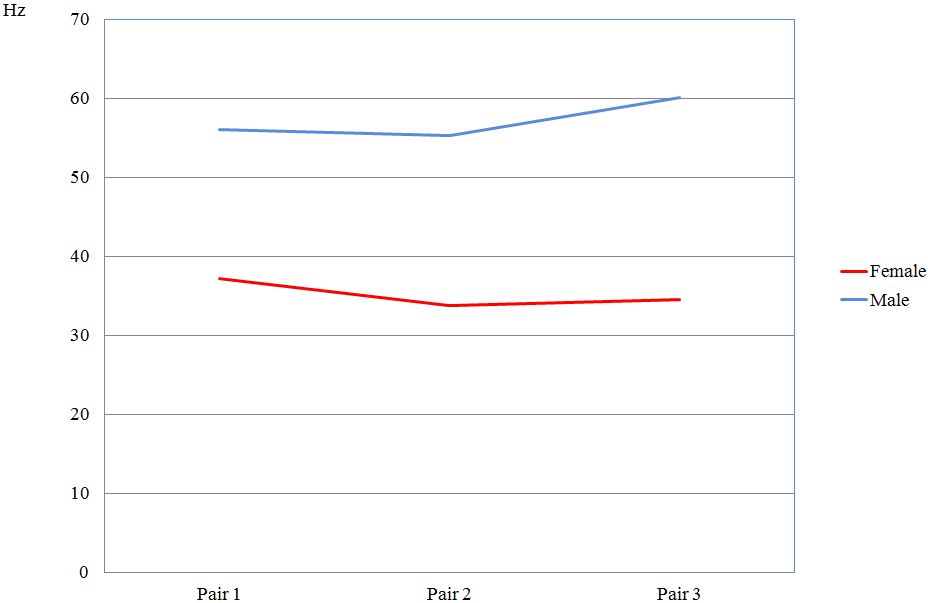
For these pairs of Common Swifts the information about the sexual affiliation is codified in the trill part at the end of the screaming call with 34 to 37 Hz for females and 55 to 60 for males (Fig. 5).M
Analysis of duetting during fighting At 6:30 pm on 12/05/2014 a third Common Swift entered the nest box. There was no recognizable reason for the Swift's entering. The conditions that afternoon were in no way extraordinary. The weather was fine with a temperature of 14 degrees Celsius. At this time the female was sitting in the nest box and at once attacked the invader directly at the entrance and fought with it. Thereupon the invader stayed in a sort of paralysis for hours. After half an hour the female left the invader alone and sat on the nest scrape. At about 7:40 pm the male of the pair returned to the nest box and did not take any notice of the invader still sitting at the entrance hole. A few minutes later the breeding pair began to concentrate on their pair grooming at the nest scrape. At 10:03 pm the female suddenly attacked the invader, which had stayed in the area of the entrance all the time and had apparently become more and more relaxed, preening its feathers. A short and intense fight between the female and the invader began, accompanied by duetting by the fighters. At about 5:30 am there was another fierce fight between the female and the invader. Finally the invader left the nest box in a rush. The spectrographic analysis indicated that the invader was a male Swift, trill part: 53Hz, s = ±3,2 , n = 38. The pitches of the individual screaming calls in the duet of the fighters were at nearly the same frequency, at 6000 Hz. The male of the breeding pair did not take any notice of the fighting activities. In contrast to this evidence Kaiser (1997) had described such fights as only occurring between Swifts of the same sex. Whether the present result represents a single incident or whether fights occur more generally between individuals of different sexes, needs to be investigated by further studies. Discussion Common Swifts are - at least to the human eye – monomorphic birds. Therefore, acoustic differences between the sexes of Common Swifts were examined in the present investigation. Van Oudheusden (2006) demonstrated in numerous investigations that the total duration of the screaming call and the total duration of the trill part showed an inter-individual significant variation, especially among Swifts from different locations. Therefore, in order to identify the sexual affiliation these parameters are excluded. Great intra- and inter-individual differences can be determined in the pitches of the screaming calls of both sexes. The present investigation shows that the pitches of the calls are highly influenced by the level of agitation of the birds and therefore they cannot be used for the sexing of Swifts. Kaiser's statement (1997) that the female Swift emits a slightly higher tone in the duet (swee) and the male a slightly deeper tone (ree), cannot be substantiated. Kaiser's conclusion may result from the method used, which was exclusively based on subjective acoustic perception. Furthermore he identified the females and males with a coloured mark at the left side or right side while duetting. Objective methods of analysis by spectrogram were not available to Kaiser. The sound impression to the human ear could have been influenced by the around 190 – 280 Hz fluctuation amplitude of the screaming part (Bretagnolle 1993). In the investigations of Bretagnolle (1993) it was already assumed that determination of the sexual affiliation is to be found in the trill part. Malacarne and Cucco (1990) also found among Pallid Swifts Apus pallidus – a circum-Mediterranean species – a typical and very constant pattern of the trills of the duetting partners of a pair. Bretagnolle already distinguished between type A (fast rhythm trill) which has a greater number of elements in the trill part and type B (slow rhythm trill). The data was insufficient to provide evidence of whether females and males show differences in the trill part. Nevertheless Bretagnolle presumed that males produced the faster trill part. In the present investigation the average of the time lapses between the single trills of the screaming calls given at the nest of three Common Swift pairs are calculated. There are significant inter-individual differences in the sequences of the trills. The intra-individual sequences of the trills are very constant. Between the two sexes the trill frequencies are clearly determined. The male of a pair always shows the faster trill frequency and therefore a higher modulation frequency in the trill part of the screaming call. When recording the screaming calls of very rapidly flying Swifts it has to be recognised that the results may be influenced by a Doppler effect. In general the following limits in the trill part can be used for the determination of the sex:
The present study shows that the determination of sexual affiliation of Common Swifts can easily be done by this method of sound recording and spectrographic analysis. References Author's address Dipl.-Biol. Konrad Ansorge, Anneröder Weg 61, D-35394 Gießen Email: mauerseglerkolonie@gmx.de © APUSlife 2015, No. 5457 ISSN 1438-2261
[StatCounter.html]
|