|
Abstract
|
| ULRICH TIGGES: | ||||||||||||||||||||||||||||||||
| Zur Brutphänologie der Mauersegler (Apus apus) - Das täglich letzte Einfliegen in Beziehung zu Umwelt- und sozialen Faktoren | ||||||||||||||||||||||||||||||||
|
Mauersegler verfolgen zwei völlig verschiedene Nächtigungsstrategien. Während Nichtbrüter in der späten Abenddämmerung hoch in den Himmel hinauffliegen, um dort zu übernachten (Weitnauer 1960), schlafen Brutvögel in ihren Nisthöhlen. Sie verbringen die Nacht nur dann im Fluge, wenn ihre Anwesenheit bei der Brutpflege nicht mehr notwendig ist oder wenn sie aus irgendeinem Grunde am Einflug gehindert werden (vgl. Lack & Lack 1952, Weitnauer 1960, Keller 1977 sowie eigene Beobachtung). Kumerloeve (1954) berichtet von nächtlichen Jagden auch in der Dunkelheit. Das abendliche Auf- und ebenso das morgendliche Herabfliegen der Mauersegler wird seit 200 Jahren beschrieben und diskutiert (vgl. Montbeillard in Weitnauer 1960, Spallanzani 1797 in Weitnauer & Scherner 1994, Guérin 1923, Weitnauer 1952, 1954, 1960). Angaben über das „Zuruhgehen“ (Schuster 1921) der Mauersegler sind in der Literatur reichlich zitiert. Sie bezeichnen das Ende der Flugbewegungen am Himmel (z.B. Hoffmann 1917, Schuster 1921, Sehlbach 1926, Haartman 1940). Mehrere Autoren setzen den letzten Einflug in Bezug zum Sonnenstand und geben die Anzahl der Minuten an, die Mauersegler über den Sonnenuntergang hinaus in der Luft bleiben:
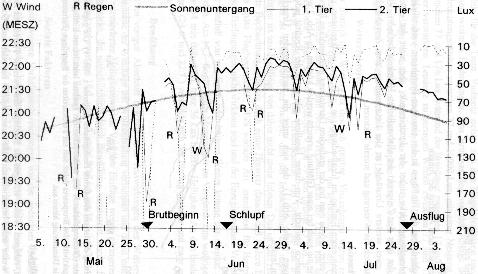
Je höher die Brutorte im Norden liegen und je länger dadurch die Dämmerung andauert, desto länger fliegen Mauersegler noch nach dem Sonnenuntergang. Trotz der Nähe zum Menschen sowie der offenen Lebensweise des Mauerseglers sind direkte Beobachtungen vom letzten Einflug selten. Quantz (1917) verzeichnete am 23. Juli zwei Einflüge um 21.45 und 21.50 Uhr in Göttingen. Swaen (1922) berichtete, dass nach seinen Beobachtungen von verschiedenen Brutplätzen in den Niederlanden Mauersegler im Juli zwischen 21.30 und 21.45 Uhr (örtlicher) Sommerzeit einfliegen, was etwa der heutigen Berliner Sommerzeit von 21.45 bis 22.00 Uhr entspricht. Keller (1977) erfasste über drei Jahre die Aktivität an einer Bruthöhle in Dresden mit Hilfe eines Automaten. Der von ihm errechnete Mittelwert des Flugendes schwankte zwischen 18.15 und 21.54 Uhr (Extreme 17.00 und 22.10 Uhr). Tigges (1994) veröffentlichte die Zeiten der letzten Einflüge während der Dauer einer Spätbrut in Berlin vom 11.5. bis 7.9. Gory (1992) registrierte in Nîmes in Südfrankreich mit Wärmemessern die Anwesenheit der Altvögel auf dem Nest. Diese Meßmethode erfasste jedoch nur die letzten Einflugzeiten des ersten Rückkehrers in denjenigen Fällen, in denen zuvor beide Alttiere ausgeflogen waren. Als einziger gibt Haartman (1949) die Helligkeit beim Zurruhegehen in der Maßeinheit Lux an. (Anm. Scheer (1949) veröffentlichte Messwerte über das Erwachen und den morgendlichen Ausflug in Darmstadt. Die von ihm errechneten Helligkeitswerte liegen zwischen 18-29 bei gutem und 103-165 Lux bei schlechtem Wetter.) Allerdings sind seine Werte nicht gemessen, sondern errechnet, was sicher den damaligen technischen Gegebenheiten anzulasten ist. Als Ergebnis seiner Untersuchung stellt er fest, dass der Mauersegler „von der Jahreszeit, geographischen Breite und Bedeckung des Himmels ziemlich unabhängig an einer bestimmten Einschlafhelligkeit“ festhält, „die etwa 4-15 Lux beträgt.“ Wird dieser Wert, den er „Schlafhelligkeit“ nennt, nicht erreicht, so folgert er weiter, würde der Mauersegler auch noch zu Mitternacht rege sein. Der Einflug bei bedecktem Himmel findet nach seinen Beobachtungen um etwa 10 Minuten früher als üblich statt, was auf der geographischen Breite seines Beobachtungsstandortes demselben Helligkeitswert entsprechen soll. Der technische Fortschritt macht es möglich, dass heute jeder preiswert in den Besitz von Luxmetern gelangen kann, weswegen die fehlenden Messreihen nun vorgelegt werden können. Zusätzlich wurden während der Beobachtungszeit die Tageshöchst- und Tagstiefsttemperaturen sowie weitere Wetterdaten der Abendstunden aufgezeichnet. Außerdem wurden sämtliche brutbiologische Daten protokolliert, um Zusammenhänge erkennen zu können. Dadurch wurde es möglich, den Einfluss sozialer Faktoren auf das Einflugverhalten zu ermitteln. 2. Material und Methode Bei den beobachteten Mauerseglern kann davon ausgegangen werden, dass es sich um ein erfahrenes Paar handelte zumindest ein 1991 beringtes Tier war wieder mit der Brut befasst (vgl. Tigges 1994). Der Brutplatz befindet sich 16 m über Grund in einem Nistkasten unter dem Fenstersturz eines vierstöckigen Altbaues in einem Gebiet mit geschlossener Bauweise der Berliner Innenstadt. Die Entfernung der Häuserfront der gegenüberliegenden Straßenseite betrug20 m. Die Kronen der vor dem Haus stehenden Linden reichen nicht bis in die Höhe des Holzkastens. Das Paar brütete ohne arteigene unmittelbare Nachbarschaft und wird durch Einflüsse, die von einer größeren Nistkolonie ausgehen, nicht beeinflusst. Ein weiteres Paar brütete wahrscheinlich auf der Rückseite der gegenüberliegenden Häuserreihe. Mit einem handelsüblichen Lichtmessgerät (Marke Lutron LX-101 Lux-Meter; laut Herstellerangaben bei 2854°K mit einer Wolfram-Halogen-Standardparallellichtlampe justiert, Messfehler ± 5%) wurde unmittelbar nach dem Einschlupf (ca. 10 bis 30 sec) die Helligkeit zwei Meter neben und ca. ein Meter unterhalb des Nistkastens gemessen. Um vergleichbare Zahlen zu erhalten, wurde mit dem Sensor des Luxmeters die an dieser Stelle jeweils größte Lichtstärke erfasst und registriert. (Zum besseren Verständnis sei hier angemerkt, dass der Wert Lux eine sehr kleine Maßeinheit ist und schon geringste Positionsänderungen des Sensors andere Werte ergeben. Aus diesem Grunde wurden die Messungen immer an derselben Stelle vorgenommen und die höchsten Werte erfasst, um diesen Unsicherheitsfaktor auszuschließen.) Die Tagestemperaturen wurden mit einem Maximum/Minimum Thermometer gemessen. Es ist etwa 90 cm neben und 40 cm unterhalb des Nistkastens im Rahmen der Balkontür angebracht. Die gemessenen Temperaturen sind bei Extremwerten jeweils um etwa 2°C gemäßigter als die vom Wetteramt veröffentlichten Daten. Sie entsprechen aber denjenigen am Nistkasten. Die Untergangszeiten der Sonne wurden vom Deutschen Wetterdienst gegen Entgelt zur Verfügung gestellt beziehen sich auf das Wetteramt im Flughafen Berlin-Tempelhof, das etwa 2700 m Luftlinie in westlicher Richtung vom Standort entfernt liegt. Das Brutpaar legte drei Eier am 26., 28. und 30. Mai, was der für dieses Gebiet bestehenden Regel entspricht. (vgl. Litzbarski in Rutschke (Ed.) 1987). Zwei Eier wurden bebrütet. Das Wetter 1995 war bis zum 23./34.6. kühl und regnerisch. Am 27.6. setzte eine stabile Periode warmen und trockenen Wetters ein, die weit über das Ende der Brutsaison hinausreichte. Da Mauersegler sich ausschließlich von Fluginsekten ernähren, sind die Wetterverhältnisse für die Reproduktion also durchweg als günstig einzustufen. An den Tagen mit Niederschlägen gab es für die Nahrungsaufnahme genügend Zeit ohne Regen. Die Beobachtungen der Flugaktivitäten an der Brutbox erfolgten regelmäßig vom Balkon oder der Wohnung aus. In der zu erwartenden Einflugzeit sowie bei ungünstigem Wetter entsprechend früher, wurde abends ununterbrochen beobachtet. In der Regel konnte ich 1995 auch die Zeit vor dem letzten Einflug kontrollieren, so dass alle kritischen Phasen, wie die Eiablage oder das Ende der Brutperiode, der letzte Einflug des zweiten Mauerseglers bis auf fünf Tage vollständig erfasst werden konnte. Vom zuerst einfliegenden Tier wurden 69 letzte Einflüge aufgezeichnet. Nachdem ich am 17. Mai um 21.45 Uhr bei 1 Lux (!) auf der Hofseite der Wohnung zwei Anflugversuche auf einen anderen Nistkasten beobachtete, habe ich, obwohl es gegen meine bisherige Erfahrung mit dem Brutpaar spricht, die Beobachtung auch nach dem von mir erwarteten letzten Einflug noch nicht beendet, sondern regelmäßig bis zur völligen Dunkelheit kontrolliert; weiterer Flugbetrieb fand bei den Brutplatzbesitzern jedoch nicht mehr statt. Das Licht hat an annähernd jeder Stelle im Raum unterschiedliche Beleuchtungsstärke. Diese ist vor der Bruthöhle aufgrund der Bebauung geringer, als im freien Himmel darüber. Mit den hier aufgelisteten Daten ist die Beleuchtungsstärke des Lichts erfasst, die in geringem Abstand von der Bruthöhle herrschte, also nicht derjenige Wert, der das Tier im Himmel veranlasste, das Nest aufzusuchen, Dieser Wert ist höher und der Vogel muss vorausberechnen, bei welcher Beleuchtungsstärke er sich entschließt, zum Übernachten in die Bruthöhle zurückzukehren, um das Einflugloch noch sicher passieren zu können. Fehleinschätzungen können vorkommen, sind jedoch offenbar äußerst gering (vgl. Weitnauer 1990). 3. Ergebnisse 1995 wurden die letzten Einflugzeiten eines brütenden Mauerseglerpaares zusammen mit der Beleuchtungsstärke sowie einigen Witterungserscheinungen und brutbiologischen Daten notiert und ausgewertet. Das Ergebnis ist in Abb. 1 dargestellt. Die Witterungsbedingungen waren gut, so dass das Verhalten der Elterntiere in dieser Brutsaison als beispielhaft gelten kann.
Abb. 1: Die letzten abendlichen Einflüge des Brutpaares, die Luxwerte beim Einflug des letzten (zweiten) Mauerseglers, der Sonnenuntergang sowie Witterungserscheinungen und Brutdaten. Frühe Heimkehr zum Nest wird durch Sozialverhalten, Brutgeschehen und Wetter bedingt Die Aufenthaltsdauer der Mauersegler lässt sich in unseren Breiten in vier Phasen unterteilen, die sich auch auf die Einflugzeiten auswirken:
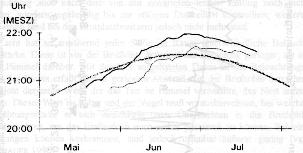
Die Unterschiede bei den Einflügen in den einzelnen Brutphasen verdeutlichen, dass die Beleuchtungsstärke sicher der dominierende, aber nicht der einzige Faktor ist, der über den Zeitpunkt der allabendlichen Heimkehr befindet. Vielmehr sind zwei weitere Umstände an der Entscheidung zum zur Ruhe gehen beteiligt, nämlich, die Witterung und soziale Faktoren. 4. Diskussion Postuliert man einen Einflug zu Idealbedingungen, z. B. bei wolkenlosem und warmen Wetter, würde man bei der grafischen Darstellung gleichmäßige Kurven erhalten, wie sie auch bei einer Glättung erreicht werden können. In Abb. 2 wird dies anschaulich gemacht. Von dieser Ideallinie gibt es in der Realität jedoch Abweichungen (siehe Abb. 1), die sich unter die nachfolgenden Ursachen subsumieren lassen.
Abb. 2: Glättung der letzten abendlichen Einflugsdaten (Legende s. Abb. 1). Die geglätteten Werte (Glättungsfaktor K 23) verdeutlichen die Tendenz beim letzten Einflug. Die Flugaktivität des Brutpaares dehnt sich mit Fortschreiten der Brutpflege (Fütterung!) in die Dämmerung hinein aus 4.1. Helligkeit Bei klarem, ruhigem Wetter erfolgen die Einflüge des zweiten Tieres mit Ausnahme der weiter unten beschriebenen Fälle ausschließlich bei oder nach Sonnenuntergang. Von den 155 beobachteten Einflügen des Brutpaares fanden 40 vor und 115 nach Sonnenuntergang statt. Der Sonnenuntergang ist aber keine fixe Größe für die Tiere. Vielmehr richtet sich ihre Flugaktivitätszeit nach verschiedenen weiteren Bedingungen. Das Brutpaar flog vor der Ablage der Eier bei Beleuchtungsstärken zwischen 77 und 10900 Lux, mit einer Neigung zu Werten zwischen 77 und 176 Lux, während der Brutdauer zwischen 17 und 360 Lux, mit einer verstärkten Tendenz zu geringen Werten zwischen 17 und 73 Lux und während der Fütterungsperiode zwischen 8 und 159 Lux, am häufigsten (35 % der Einflüge) zwischen 17 und 20 Lux ein. An den Ausflugtagen der Küken kehrten beide Elterntiere nicht zum Brutplatz zurück. Nach dem Ausfliegen der Jungen lagen die Werte zwischen 9 und 18 Lux. 4.1.1. Wetterfaktoren Der Einfluss des Wetters ist deutlich spürbar. Vierzehnmal waren regnerisches Wetter, Regen oder Gewitter (in Abb. 1 alle mit R bezeichnet) die Ursache für die vorzeitige Rückkehr zum Nest. Vergleicht man die Zeiten mit solchen von regenlosen Tagen, lag sie zwischen 9 und 59 min (Mittel 25 min, n=14) früher, und zwar unabhängig von der Lichtintensität. Ein Grund für das frühere Einfliegen ist sicher darin zu suchen, dass der Insektenflug bei nassem Wetter geringer ist und Jagdflüge von daher nicht mehr lohnen. Das ist eine recht interessante Beobachtung, gibt sie doch einen Hinweis darauf, dass andere Impulse für eine späte Flugbereitschaft, etwa sozialer Art, zumindest für Brutpaare unter diesen Bedingungen nicht wirksam sind und der Energiehaushalt Vorrang besitzt. Der Einfluss von windigem oder böigem Wetter konnte nur dreimal festgestellt werden. Am 14. 7. flogen beide Mauersegler im Vergleich zum wolkigen 17. 7. 67 bzw. 54 min früher ein, trotz 36 Lux höherer Helligkeit. An den beiden anderen Tagen (24.6. und 22.7.) verfrühte sich der Einflug geringer und zwar um etwa 10 min. 4.1.2. Bewölkung Die im vorigen Abschnitt beschriebenen Auswirkungen des Wetters sind mit Ausnahme der Windtage mit Wolkenbildung verbunden und damit natürlich auch mit verminderten Helligkeitswerten. Verfrühte Einflüge wegen geringerer Beleuchtungsstärke, die ausschließlich durch Bewölkung verursacht wurde, fanden nur zweimal und zwar am 16.5. und 6. 7. statt. Weil ich am 16. Mai um 20.34 Uhr noch nicht mit dem letzten Einflug rechnete, habe ich versäumt, an diesem Tag die Luxwerte zu messen. 4.1.3. Soziale Faktoren Für neun weitere Verhaltensabweichungen von der gedachten Ideallinie gibt es keine physikalischen Erklärungen. Das ist am 5. und 19.5. bei heiterem Wetter (1050 bzw. 374 Lux), am 21., 22. und 23.5. bei blauem Himmel, am 25. und 27.5. (wolkenlos, 10900 Lux bzw. bedeckt und 4240 Lux) sowie am 2. und 4.7. (jeweils bedeckt, 36 und 20 Lux) der Fall. Die Temperaturwerte an diesen Tagen lagen beim Minimum zwischen 8 und 19°C, beim Maximum zwischen 18 und 28°C, zeigten also ebenfalls keine Auffälligkeit. Als Erklärungen bieten sich außer beim 5.5., bei dem es sich um den Ankunftstag des ersten Tieres handelte, soziale Faktoren an. Am 26. und 28.5. erfolgte die Eiablage, die dem frühen Einfliegen an den Vorabenden einen Sinn gibt. Natürlich geht diese Beobachtung von zwei Daten nicht über den statistischen Zufall hinaus. Es ist aber interessant, dass dieses Verhalten vor beiden Legedaten stattfand und von beiden Partnern gezeigt wurde. Der Einflug am 25.5. fand überdies gemeinsam statt. Am Vorabend der Ablage des dritten Eies am 30.5. herrschte schlechtes Wetter, so dass er nicht als Beweis herangezogen werden kann. Bei allen fünf Brutpaaren, deren Anwesenheit Gory (1992) protokollierte, ist bei am Morgen gelegten Eiern für den Vorabend ebenfalls ein früheres Einfliegen (des ersten Tieres) festzustellen. Für die These von sozialen Faktoren, die eine frühe Rückkehr veranlassen, sprechen auch die folgenden Ereignisse. Der 21.5. war der 5. Tag vor der ersten Eiablage. Bei Mauerseglern ist diese Frist die Zeitdauer der Eibildung (vgl. Lack 1956, Weitnauer & Scherner in Glutz von Blotzheim 1994), so dass an den entsprechenden Abenden sexuelles Verhalten angenommen werden kann. Für den 19.5. sowie die Tage danach gilt das gleiche. In den Grafiken bei Gory ist ebenfalls einmal ein abendlicher vorzeitiger Einflug ins Nest am fünften Tag vor der Eiablage dargestellt. Die Termine am 2. und 4.7. schließlich liegen einen Tag vor der Augenöffnung der Nestlinge (vgl. Vizyová & Janiga 1986), beide Male reagierten die Elterntiere auf dieses Ereignis. Weil dieses Geschehen in der Zeitspanne des permanenten Huderns eintritt, konnte es von der Messmethode Gorys nicht erfasst werden. Das Einflugverhalten beider Mauersegler stimmt in hohem Maße überein. An zehn Tagen flogen sie unmittelbar zusammen in die Nistbox, z.T. mit einer Geschwindigkeit, dass der Einflug innerhalb des Bruchteils einer Sekunde stattfand. Manchmal konnte ich beobachten, wie ein Tier den Flug zusätzlich abbremsen musste, um zunächst seinen Partner einschlüpfen zu lassen. Die Verfolgungsentfernung ist in solchen Fällen wohl noch dichter als bei dem zuvor beschriebenen Fall, was mit dem menschlichen Auge aber nicht mehr erfasst werden kann. An 13 Tagen kehrten beide Tiere innerhalb von zwei Minuten vom letzten Ausflug zurück. Die größte Synchronie herrschte in der Zeit ohne Brutpflichten, also vor dem Brutbeginn und nach einem Termin, der vier Tage vor dem Ausflug der Jungen lag. Etwa zu dieser Zeit kann die Phase der Jungenfütterung bei den Mauerseglern bereits beendet sein (vgl. hierzu Angaben bei Schulz 1881, Weitnauer 1947, Lack & Lack 1951, Wendt 1988, Polaschek 1992). Nach der Eiablage und dem 11. Tag nach dem Schlupf flogen beide Elterntiere nur einmal zusammen ein. Diese Phase ist die des ständigen Huderns (vgl. Cramp 1994). Es ist also festzustellen, dass die Mauersegler über einen beeindruckenden sozialen Kontakt verfügen. Interessant wäre in diesem Zusammenhang natürlich die Antwort auf die Frage der Rolle der Geschlechter beim letzten Einflug. Zwar wird bei Angaben zur Brutbiologie gelegentlich auf ein Geschlecht besonders hingewiesen (z.B. Weitnauer 1947, Flath 1980, Bezzel 1985), sofern jedoch nicht dargestellt wird, wie die Geschlechter identifiziert und weiterhin erkannt worden sind, sind solche Bemerkungen wertlos, da Männchen und Weibchen ohne Sektion nur bei der Kopulation, und, wie Kaiser (1997) jetzt entdeckte, beim Rufen eindeutig identifiziert werden können. Um diese Frage zu beantworten, müssten die Tiere auf eine Art und Weise gekennzeichnet werden, die eine Saison lang ohne weitere Fänge sichtbar bliebe, was der Bundesartenschutzverordnung unterfallen würde. Bei den hier vorgestellten Untersuchungen wurde das Geschlecht der Vögel nicht identifiziert, sondern sie wurden nach der Reihenfolge des Einfliegens nummeriert. Der letzte Heimkehrer ins Nest kann also sowohl das Weibchen als auch das Männchen sein. Bisher ist bei Mauerseglern gänzlich unbekannt, ob - abgesehen von der Kopulation und der Stimmlage - überhaupt ein Verhalten geschlechtsspezifisch ausgeprägt ist, das betrifft natürlich auch die abendliche Reihenfolge der Rückkehr ins Nest. 5. Zusammenfassung Von einem Mauerseglerpaar, das unter günstigen Wetterbedingungen erfolgreich seine Brut aufziehen konnte, wurden während der gesamten Aufenthaltsdauer am Nistort die letzten abendlichen Einflüge registriert. Sie wurden mit dem Sonnenuntergang verglichen und sowohl mit Witterungsbedingungen, als auch brutbiologischen Daten in Beziehung gesetzt. Es wurde festgestellt, dass der Sonnenuntergang nur eine relative Größe für die Impulse zum abendlichen letzten Einflug der Mauersegler ist. Bezugsgrößen sind die Helligkeit in Abhängigkeit von Witterungserscheinungen sowie soziale Faktoren. Die Bedeutung der sozialen Faktoren muss durch weitere Beobachtungen anderer Brutpaare überprüft werden, wozu an dieser Stelle aufgerufen werden soll. Ich danke Dr. Henry Hahnke, vormals Museum für Naturkunde zu Berlin, für die Diskussionen.
|
||||||||||||||||||||||||||||||||
|
(Printed
1999 in Berliner ornithologischer Bericht. Republished with permission.)
|
||||||||||||||||||||||||||||||||
| © APUSlife
2000, No. 0067
ISSN 1438-2261 |
||||||||||||||||||||||||||||||||